
遺伝子診断技術(1)
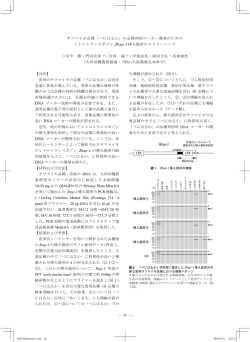
植物医科学応用実験 II 第1回 第7回 遺伝子診断技術 テキスト 目 次 i) 核酸抽出 2 ii ) PCR 5 iii ) アガロースゲル電気泳動 7 iv ) シークエンス反応 11 v) シーケンス結果の解析 13 1 植物病原体の種々の診断法の中で、遺伝子診断法は近年めざましく技術が発展しつつあ る。本実習では、その中で特に基礎的な方法を学ぶ。具体的には、分離された菌体から DNA を抽出し、ユニバーサルプライマーを用いて rDNA 遺伝子の ITS 領域を PCR によ り増幅する。電気泳動による増幅産物のチェック、シークエンス反応を行って、その塩基 配列を決定し、GenBank に登録された塩基配列に対して相同性検索をかけることにより、 その属種を推定する。 i) 核酸抽出 ①50 ml ビーカーの口に二重にしたガーゼを被せ、輪ゴムで固定する。 ②ガーゼの上に液体培養した菌体をのせ、薬さじでこして水気を切る。 上から milliQ 水を注いで培地を洗い流し、薬さじでこす。 ③菌体をろ紙の上に載せて更に水気を切り、ピンセットを使って 5 mm 角程度ちぎる。 ④菌体を乳鉢に入れ、軽く乳棒で延ばす。 ⑤乳鉢に液体窒素を注ぎ、粉末になるまでよくすり潰す。 (液体窒素使用の際は軍手をしてその上にサニメント手袋を着用すること) ⑥DNA 抽出バッファー 1 ml を乳鉢に加え、乳棒でさらによく混合する。 ⑦磨砕液 500 µl を 1.5ml チューブに取る。 (チップの先端をあらかじめハサミで切っておく) ⑧チューブに取った磨砕液に 500 µl のフェノール・クロロホルム(フェノクロ)を 加える。 (サニメント手袋、保護メガネを着用のこと) (フェノクロの取り方:上層は平衡化のための Tris バッファーであり、下層が目的のフ ェノクロである。いくつかの有機溶媒はマイクロピペットの中で揮発して圧力を上げ、 チップ先端から漏れることがある。フェノクロもそのような試薬であるため、取る際 には、下層にチップの先端を差し込んだ後、何度かゆっくりとピペッティングしてピ ペットの中を有機溶媒の蒸気で満たして安定させてから所定量を吸う。) ⑨液をボルテクスミキサーで約1分よく混合する。 (もれないようにフタをよく閉めておくこと) ⑩15,000 rpm(25℃)で 5 分間遠心する。ローターにチューブをセットし遠心する際は、 必ずバランスを取ること。 (偶数本であればそれぞれ向かい合わせに、奇数本であれば 2 3本を均等に配置し残りを向かい合わせにセットする) ⑪遠心後、チューブ内の液は下層の有機溶媒層(フェノクロ)、上層の水層(TE バッフ ァー)、および中間層(変性タンパク質等)に分離する(図参照)。上層の水層に DNA が溶解しているため、水層を別のチューブに回収する(約 400μl)。 (中間層を取らないように注意する。タンパク質が混入すると、PCR 反応等の今後の 実験の阻害要因になる。水層のすべてを取る必要はない。) ⑫回収した水層に等量( µl)のクロロホルム・イソアミルアルコール(クロイソ) を加える。 (クロイソを取る際も、上述の注意点と同様ピペッティングして安定させてから吸う) ⑬液をボルテクスミキサーでよく混合し、15,000 rpm(25℃)で 5 分間遠心する。 ⑭遠心後、分離した上層の水層を別のチューブに回収する(図参照)。 ⑮回収した液量の 1/10 量( µl)の 3 M 酢酸ナトリウム水溶液を加える。次いで、回 収した液量の 2.5 倍量( µl)の 99%エタノールを加え、転倒混和する。 ⑯15,000 rpm(4℃)で 10 分間遠心する。 ⑰チューブの底に DNA が沈澱している(ペレット;図参照)。 デカンテーションにより、上清を捨てる。 ⑱70%エタノールを 500 µl 加え、そのまま 15,000 rpm(4℃)で 5 分間遠心する。 ⑲デカンテーションにより上清を捨て、チビタン(小型卓上遠心機)を用いて残液を一 度落としてから 10 µl のマイクロピペッターで丁寧に取り除く。この際ペレットを突い て崩さないように注意すること。 ⑳チューブの蓋をあけたまま、キムワイプの上に逆さに置いて 10 分間風乾する。 (上清の除去が十分でないと乾燥に時間がかかる。)(第 1 回ここまで) 21. 20 µl の TE バッファーを加え、ペレットを溶解する。 ボルテックスでよく撹拌し、尖端を切ったチップでピペッティングする。 ポイント フェノクロ抽出、クロイソ抽出の際に中間層を吸い込むことなく作業ができた か。エタノール沈殿後のペレットの様子はどのようであったか。ペレットを失うことなく 作業ができたか。 3 課 題 ①クロロホルムの MSDS(化学物質安全性データシート)をインターネットで調べ、有害 性と取り扱いの方法について、簡単に実験ノートにまとめよ。 ②DNA のエタノール沈殿について、その原理を調べて記せ。 核酸の抽出について 生物体には核酸以外に、多くのタンパク質などが含まれているため、単純にバッファーで磨 砕しただけでは残ったタンパク質などが核酸を分解したり、PCR 反応等を阻害することがあ る。このため、タンパク質を変性させるフェノールを加えて遠心分離することで、タンパク質 を不溶化し、中間層に集める(フェノクロ処理)。核酸は水溶性分子であるため、上層の水層に 溶解する。しかし、フェノールは前述の通りタンパク質を変性させるため、微量でも水層中に 残存すると、後の PCR 反応で使用する酵素まで変性させてしまう。このため、クロロホルムを 用いてフェノールを回収し、取り除く操作が必要となる(クロイソ処理)。こうして水層に集め た核酸は、高濃度の塩(今回は 3M 酢酸ナトリウム)とエタノールの存在下で沈澱させる(コロ イドの塩析、エタノール沈澱)。最後に残った塩を 70% エタノール(の中の水分)に回収し、 核酸を精製する。 参考図 水層 中間層 有機溶媒層 フェノクロ後の写真( i-⑪)。 クロイソ後の写真( i-⑭)。 水層の液を取る。 水層の液を取る。 ペレット エタノール沈殿後の写真(i-⑰)。不純物が少ない場合はペレットの色は薄くなる。 4 ii ) PCR ①発泡スチロールに氷を詰めておく。 PCR 用試薬(ExTaq 以外)を解かして良く混ぜ、氷にさしておく。 ②プレミックスを作る(各班1本:7 or 8 サンプル分(プレミックスは余裕をみてその+1 サンプル分作製する))。 今回、班の人数分(5 or 6)のサンプルに加え、ポジティブコントロール(PC; テンプ レート DNA としてすでに増幅されることが確認されている抽出 DNA サンプルを使用す る)とネガティブコントロール(NC; テンプレート DNA として DDW(milliQ 水)を使用 する)の計 7 or 8 サンプルの反応を行う。 PCR 用試薬(ExTaq 以外)を 1.5 ml チューブに加えてボルテックスでよく撹拌した 後、ExTaq を加えて(※1)、ピペッティングにてよく混合する(酵素はグリセロール 溶液に溶解しているため、比重が大きい。従って、よく混合しないと酵素が均一になら ず、反応が失敗するので特に注意する。酵素を添加する際にチューブを注視すれば、界 面が観察されるはずである。これがなくなるまでよく混ぜる)。 1 sample DDW(milliQ 水) 8 or 9(プレミックス) 13.8 µl µl 2.5 mM dNTP mix. 2.5 µl µl 10 PCR Buffer 2.5 µl µl 5 µM ITS1 プライマー 2.5 µl µl 5 µM ITS4 プライマー 2.5 µl µl ExTaq(※1) 0.2 µl µl subtotal 24 µl / sample テンプレート DNA total ※1 1 µl (プレミックスを分注後) 25 µl / sample ExTaq(タカラバイオ製)は DNA ポリメラーゼ(つまり酵素)である。酵素類 は室温で置いておくと失活するため、使う直前にフリーザーから出すようにする。使用 中はアイスボックスに挿しておき、使用後すぐにフリーザーに戻す。今回は各実験室の 教卓のところに置いておくので、各班 1.5 ml チューブを氷に挿した状態で持参し、所定 量をとって入れる。また、酵素は空気に触れても失活するため、添加後は泡立てないよ うにする(ボルテクスミキサーでなく、ピペッティング(中サイズのピペッターを使用 する)にて混合する)。容量の少ない液体をチューブに入れる際には液面に接触させて添 加すること。 ③PCR チューブの蓋の部分に班番号とサンプル名を書いておく(チューブの側面などに 書くと PCR 反応時に消えることがあるため避ける)。 5 ④プレミックスを 7 or 8 本の PCR チューブに 24 µl ずつ分注する。 ⑤テンプレートとなる DNA を 10 倍希釈しておく。新しい 1.5 ml チューブに DDW(milliQ 水)を 9 µl、サンプル DNA を 1 µl 加えて希釈する。各 PCR チューブに希釈した DNA を 1 µl 入れる。(PC にはコントロール DNA(これはあらかじめ希釈したものを配布するので、希 釈せずにそのまま使う)を、NC には DDW(milliQ 水)を 1 µl 入れる。) ⑥PCR チューブを Veriti サーマルサイクラー(Applied Biosystems 社)用のアダプタ ーにセットした後、機械にセットしてフタをする。 ⑦サーマルサイクラーに以下の条件を入力し、PCR 反応を行う。 96℃ 5:00 96℃ 0:40 52℃ 0:40 35 72℃ 1:00 72℃ 7:00 4℃ ∞ PCR (Polymerase Chain Reaction) 日本語ではポリメラーゼ連鎖反応と呼ぶ。2 つのプライマーと呼ばれる、鋳型 DNA に 相補的な短い一本鎖 DNA 間の配列を特異的に増幅する技術である。 鋳型 DNA 二本鎖の変性 (96℃)、プライマーの鋳型 DNA とのアニーリング(今回は 52℃。 プライマーおよび反応性によって数℃ほど調節する)、DNA 伸長(今回は 72℃。DNA ポ リメラーゼによって至適温度が決まっている)の 3 つの温度を 1 サイクルとし、このサイ クルを 25~35 回ほど繰り返すことで指数関数的に DNA を増幅させる(理論上は 1 サイク ルで 2 倍に増幅する)。 ITS 領域 今回増幅する DNA はリボソーム DNA の ITS(Internal Transcribed Spacer)領域で ある。用いるプライマーは Forward primer として ITS1 と Reverse primer として ITS4 である。これらのプライマーは真核生物に共通した配列をもとに設計されたプライマー (ユニバーサルプライマー)であるため、菌類のほか、動植物についても増幅することが できるが、原核生物である細菌については増幅することはできない。両者に挟まれる領域 の長さは生物によっても異なるがおおむね 0.6 kb 程度であるため、PCR 後のアガロース ゲル電気泳動により想定するサイズのバンドが検出されれば、以後のシークエンス解析に 持ち込むことができる。 Forward primer: ITS1 (5´-TCCGTAGGTGAACCTGCGG-3´) Reverse primer: (5´-TCCTCCGCTTATTGATATGC-3´) ITS4 6 ポイント プレミックス作成の際にきちんとピペッティングにて混合ができたか。また酵 素を失活させないための注意事項が守れたか確認する。 課 題 ①今回の場合のポジティブおよびネガティブコントロールの意義をそれぞれ考えて実験ノ ートに記せ。どういった結果になった場合に問題解決のため役立つのか。 ②プレミックスを作製する理由は省力のためであるが、別の利点は何か。 ③PCR の原理について、模式図を描いて説明せよ。 iii ) アガロースゲル電気泳動 事前準備 ① 1 TAE バッファーを作製する。 1 L のメディウム瓶に以下の通り試薬を加え混合する。 DDW(milliQ 水) 900 ml トリス(Tris (hydroxymethyl) aminomethane) 4.84 g 1,145 µl 酢酸 0.5 M EDTA (pH 8.0) 2 ml DDW(milliQ 水) Up to 1,000 ml Total 1,000 ml ② 0.7 % アガロースゲルを作製する。 Mupid-Ace (アドバンス社)のゲルメーカーを用いる(図参照)。 電気泳動用アガロース 0.21 g を計り取り、200 ml の三角フラスコに入れる。 1 TAE バッファーを 30 ml 加える。(0.7%) フラスコの口にゆるくラップをかけ、電子レンジでアガロースを溶解する。 (中身が沸騰したら止めることを 3、4 回繰り返す。) 軍手をし、その上からサニメント手袋をした状態で、電子レンジからフラスコを取り 出す。(この際に突沸するので注意!)アガロースの粒が残っていないことを確認。 振り混ぜて冷ます(かろうじて素手で持てる程度まで)。 7 ゲルトレイ台にゲルトレイをセットし、ゲルを流し込む。 チップの口の広い側でゲル板を抑え、ゲルトレイの下の気泡を除く。 ゲルトレイの一番上の溝にコーム(細い側を下に)を挿す(図参照)。 ホコリが入らないようラップをかけ、静置する。(約 30 分) 冷えて固まったら、穴(ウェル)が崩れないようコームを慎重に抜く。 1 TAE バッファーを適量入れたタイトボックスにゲルトレイごとゲルを入れて次回 まで保存する。(第 2 回ここまで) 参考図 ゲルメーカーの構成。 コーム(左)、ゲルトレイ台(右上)、ゲルトレイ(右下)。 ゲルメーカーの組立図。 アガロースゲル電気泳動 核酸は一般に負に帯電している分子であることから、核酸を含む溶液に電荷をかけると 陰極側から陽極側に移動する。このときアガロース等のゲルを用いることで、分子量の大 きい核酸は遅く、分子量の小さい核酸は速くゲル内を移動する。このため、核酸の分子量 によってゲル内の移動距離が分かれることになる。 ゲルによる泳動後の核酸は適当な染色法により検出する。エチジウムブロマイドは DNA 二本鎖の間に入り込む(インターカレート)と、紫外線照射下で蛍光を発する物質で、こ れで染色することでゲル内の核酸がバンドとして検出される。 8 電気泳動手順 ③作製したゲルをゲルトレイごと泳動槽(Mupid-Ace)にセットし、ゲルがひたるまで、 1 TAE バッファーを注ぐ。また、200 µl のピペットでウェルを洗浄しておくと良い。 ④PCR チューブに入った PCR サンプルを用意する。 (前回の実習後にスタッフで回収済み) ⑤パラフィルムを机の上に置き、その上に loading dye 5 µl をサンプルの数だけ載せる。 順に PCR 反応産物 1 µl を dye に加えつつピペッティングして混合する。 ⑥混合したサンプル(全量 6 µl)をアガロースゲルのウェル(穴)にロードする。 λHind Ⅲマーカーは 4 µl ロードする。ロードするサンプル順を記録しておくこと。 ゲルは 17 レーンあるので、左右は空けて中央付近の 8 レーンを使用する。 ローディングの際はチップの先に空気が残っていると液中で空気が上がって一緒に DNA も流れていってしまうので、先端の空気を除いて行うと良い。また、左手をピペッ トの先に添えると安定してローディングができる。ピペットの向きはウェルの縦方向で はなく、横方向に向けると、ゲルを突き刺す心配なくローディングしやすい。 ⑦85V で 30 分間泳動する。 ⑧染色バッファーを作製する。 タイトボックスに約 150 ml の 1 TAE バッファーを加え、エチジウムブロマイド溶液 (EtBr)を 10 µl 加えて振り混ぜる。 ※EtBr は変異原性があるため取扱いに気をつけること。必ずサニメント手袋、保護メガ ネを着用して作業する。万一皮膚に付着した場合にはすぐに大量の水で洗い流す。 ⑨泳動後のゲルを泳動槽から取り出し、ゲルトレイからゲルを外しながら(指で軽く押せ ば良い)、染色バッファーの入ったタイトボックスにゲルを入れて 10 分間静置して染色 する。 ⑩染色したゲルをトランスイルミネーター(Mupid Scope-WD)で確認、写真撮影する。 (サニメント手袋、保護メガネを着用) ビニールを実験台に敷き、ゲルをヘラで取り出して気泡が入らないように載せる。 Mupid Scope-WD の本体フタをあけ、ゲルをビニールごと持って UV 照射部に載せる。 シートの下にも気泡が入らないようにする。 本体フタをとじ、電源を入れて UV 照射する。上部窓から覗いて泳動バンドを確認する。 上部窓にデジカメを装着し、撮影する(マクロモードにし、ズームアウトする)。 ※UV を直視すると目を痛めるため、本体フタを開けた状態で UV 電源を入れないよう に注意すること。 (第 3 回ここまで) 9 電気泳動像の見方 電気泳動を行うと、すでに大きさと量が分かっている DNA(今回はλHindIII マーカ ー)と比較することにより、目的のサンプルの大きさと量が分かる。 λHindIII マーカーは下図のような泳動パターンとなり、肉眼では7本のバンドが見ら れるはずであり、それぞれは右に記したような大きさの DNA 断片である。今回は、およ そ 0.7 kb の遺伝子領域を増幅しようとしているので、もし増幅が成功すれば、564 bp の バンドよりもやや上の位置にバンドが見られるはずである。 また、その増幅された DNA の量も推定することが可能である。今回はλHindIII マーカ ーを 4 µl 泳動すると、左のような DNA 量となるように調整してある。したがって、仮に サンプル 1 µl 泳動して 2 kbp のバンドと同じくらいの明るさのバンドであった場合、原液 は 10 ng/µl であると推定することができる。 ポイント もれなくローディングはできたか。穴あきのゲルになっていないか(ローディ ング後に Dye が下に流れ落ちる)。染色はされているか(マーカーがきちんと光っている かどうかで判断できる)。 課 題 写真データは実験ノートに貼付け、各レーンの説明(サンプル名)を記載する。 泳動像から、自分および班員の増幅 DNA サンプルのおおよそ大きさと量を推定する。菌 の種類によって ITS 領域の大きさに差があるかなどに注目する。また、失敗した場合は PC や NC などの結果も考慮して考察する。 10 iv ) シークエンス反応 ①精製処理(ExoStar 反応):PCR 増幅産物 5 µl、エキソヌクレアーゼ 1 µl、アルカリフ ォスファターゼ 1 µl を PCR チューブに混合し、サーマルサイクラーにて以下の条件で 酵素処理を行う。 37℃ 15:00 80℃ 15:00 4℃ ∞ PCR 増幅産物処理 PCR 増副産物をベクターにクローニングせずにシークエンス反応にかける(ダイレクト シークエンス)ためには精製処理を行わなければならない。これは PCR 未反応のプライマ ーや dNTP が、シークエンス反応に悪影響を与えるためである。 精製処理の方法としては、ゲル精製や限外ろ過法など様々なものがあるが、今回実習で 行うのは illustra ExoStar(GE ヘルスケア社製)という試薬を用いて、プライマーと dNTP を不活化する極めて簡便な方法である。 illustra ExoStar の中には、エキソヌクレアーゼ I とアルカリフォスファターゼの2種の 酵素が入っており、前者がプライマーなどの1本鎖 DNA を消化し、後者が dNTP のリン 酸基を除去することで、両者がシークエンス反応を阻害しないような形とする。この酵素 反応は 37℃で行い、80℃で酵素を失活させることにより、これらの試薬自身が後のシーク エンス反応を阻害しないようにする。 ②シークエンス反応:以下の DDW からプライマーまでの試薬をよく混合し(1.5 ml チュ ーブにプレミックスを作る)、BigDye を加えてピペッティングしたものを PCR チュー ブに分注する(プレミックスは余裕をみて班の人数分+1 倍量調整する)。各 PCR チュー ブに精製処理したサンプルを加える。 1 sample 6 or 7 DDW(milliQ 水) 3.75 µl µl 5 x Sequence Buffer 1.75 µl µl 3.2 µM ITS1 プライマー 0.5 µl µl BigDye Terminator v3.1(※) 0.5 µl µl Template(ExoStar 反応産物) 3.5 µl 分注後 Total 10 µl ※前述 PCR の項の酵素類使用の際の注意点(室温に温めないこと、ピペッティングする こと等)を守ること。 11 ③PCR チューブをサーマルサイクラーにセットし、以下の条件でシークエンス反応を行う。 96℃ 3:00 96℃ 0:15 50℃ 0:05 25 60℃ 4:00 4℃ ∞ (第 5 回ここまで) サンガー法による塩基配列の決定 今回行う塩基配列決定の手法はサンガー法(ダイデオキシ法)と呼ばれるものであり、 その原理の概要は次の通りである。 まず、DNA ポリメラーゼを用いて目的とする DNA の相補鎖を合成するが、その際に DNA 合成の原料である dNTP (deoxynucleotide triphosphate)に加えて ddNTP (dideoxynucleotide triphosphate)という分子を少量加えておく。ddNTP とは dNTP の 3´の位置の -OH 基が-H 基になったもので、DNA 合成の際にこれが取り込まれると、その後の DNA 鎖の伸長がストップする。こうして DNA 合成反応後は、合成がストップしたいろいろの長 さの合成産物ができるが、このとき、例えば ddATP が取り込まれて反応がストップした産 物の末端塩基は全て A である、と言うことができる。同様に ddTTP, ddGTP, ddCTP につ いても同じことがいえるが、今回用いるシークエンス反応キット(BigDye Terminator: Applied Biosystems 社製)は、それぞれの ddNTP が違う種類の蛍光色素で修飾されてお り、得られた合成産物を電気泳動してその長さにより分離しつつ、レーザー光をあてて蛍 光色素を検出する(3130xl ジェネティックアナライザ:Applied Biosystems 社製)ことで、 塩基配列を決定することができる。 ④膨潤済みレジンをよくボルテックスして懸濁し、カラムチューブに 750μl 充填する。 ⑤カラムチューブを、排水用 2ml チューブにセットして 750 g で3分間遠心を行い、カ ラムの余分なバッファーを除く。 ⑥カラムチューブを、精製産物回収用の新しい 1.5ml チューブにセットする。シークエン スリアクション産物に DDW 10μl を加え(合計で 20μl になる)、全量をカラムに染み 込ませる。(この時、サンプルが確実にカラムレジンに染み込む様に注意する。) ⑦750 g で3分間遠心を行い、精製 DNA 溶液を回収する。 ⑧チューブに回収した DNA 溶液を 96 穴シークエンス用プレートの各班の穴に移しかえる。 ※班ごとに入れる場所を指定するので、絶対に間違えないこと。 ⑨シーケンサーにかける。 12 ポイント 「酵素類使用の際の注意点」を守ることが出来たか。カラムに添加する際に、 壁面ではなく、カラム担体にしみ込ませられたか。 課 題 サンガー法について調べ、手書きの図を用いて実験ノートに記せ。 カラム精製 シーケンス反応後の産物には未反応の ddNTP などがあり、シーケンサー内での電気泳 動に悪影響を与えるので、目的の反応産物のみを得るため精製する必要がある。精製の方 法としては、エタノール沈殿などの方法もあるが、今回は一般的に用いられるカラム精製 により行う。 ここで言うカラムとはカラムクロマトグラフィーのことで、クロマトグラフィーの固定 相が円柱形(カラム)をなすことからこのように呼ばれる。クロマトグラフィーは多成分 の混合物を分離する方法で、今回はその固定相に FastGene G50(ニッポンジーン社)と いうゲルろ過用担体を用いる。FastGene G50 に載せられた反応混合物のうち、分子量の 比較的小さな未反応 ddNTP は担体粒子の網目内にトラップされて残る一方、分子量の大 きな目的反応産物は粒子間の空隙から流れ落ちて出てくる。 v ) シーケンス結果の解析 ①シーケンス結果のデータを各自の PC に移す。 (データはテキストファイルの形式で教卓の PC の中にある) ②NCBI の BLAST を開く。 http://blast.ncbi.nlm.nih.gov/Blast.cgi ③ Basic BLAST 項目内の nucleotide-blast を選択。 ④最上段(Enter Query Sequence)の「Enter accession number, gi, or FASTA sequence」 のボックスに調べたい配列をペーストする。 ④中段(Choose Search Set)の Database 欄の「Others (nr etc.)」にチェックを入れる。 ⑤最下段の「BLAST」を押す。 ⑥出てきた検索結果を元に考察を行う。 13 課 題 A E のうち、班で解析した全てのサンプルについて、以下のことをまとめよ。 ①上位数十件の検索結果にはどのような生物の配列が認められたか列挙せよ(属名/種名 とその相同性を挙げる)。このことから、A E の菌類は何であったと推定されるか、数 字等の根拠を用いて考察せよ(菌類の有性世代、無性世代の名称に注意する→必要に応 じて調べる)。今回の解析で属名のみ明らかになったのか、あるいは種名まで明らかであ るのか、考察する。 ②上手く読めなかったサンプルに関して、どこがいけなかったのか、サンプルと作業手順 (とくに電気泳動後の結果)を振り返って照らし合わせることにより考察せよ。 シークエンスデータの見方 前述の通り、シーケンサーはシークエンス反応産物をキャピラリー内で電気泳動しつ つ、A, T, G, C の塩基に対応する4色の蛍光色素のシグナルを読み取るものである。その 結果、シークエンスデータは、横軸を反応産物の分子量、縦軸を蛍光シグナルの強度とし た4本の(色の異なる)波形データの形で出力される(図参照)。そのデータをもとに、 シークエンス解析ソフトで塩基を読みとって(ベースコーリング)テキストデータに変換 する(図参照)。 波形データにおいて単一の塩基のピークが認められれば、ソフトウェアの解析でその塩 基を確実に決めることができるが、非特異反応が多いときなど、何らかの理由で波形デー タが重なり合う場合にはその塩基が特定できず、”N”などの表示となる。今回のプロトコ ルによる 3130xl シーケンサーにおける配列解析は 600-700 bp 程度が正確に解読できる限 度なので、テキストデータのうち前後の末端で N の多い領域はカットして BLAST 検索に 用いる。 図 波形データとベースコールされた塩基配列 14 BLAST 検索結果の見方 Query:自分が送信した配列 Subject:検索してヒットした配列 1. 一番上に Graphic Summary が表示される。 横軸は Query の配列の全長を示す。バーの一本一本が検索された Subject を表し、 Query のどの領域にヒットしたかが一目で分かるようになっている。バーの色はアライ メントスコアを示しており、赤色が最も高得点である。マウスポインタをバーに合わせ ると、上部窓に Description が表示され、クリック先は Alignment にリンクされている。 2. 次に One-line Descriptions が表示される。 ヒットした Subject がスコア順に一行表示され、以下のデータが示される。 i) Accession:Subject の遺伝子のアクセッションナンバー (GenBank のデータにリンクしている) ii) Description:遺伝子の説明 iii) Max score, Total score:アライメントがどの程度マッチしているかの得点 (Alignment にリンクしている) iv) Query coverage:アライメントされた配列が Query のどの程度をカバーする かの割合 v) E value:そのスコアの配列が偶然にアライメントされてしまう数の期待値 (数値が小さいほど、偶然ではないことを示している) vi) Ident:相同性 3. 一番下に詳細な Alignments が表示される。 (アクセッションナンバーが GenBank のデータにリンクしている) 上記 1, 2, 3 は表示の仕方が異なるだけで、すべて同じ結果を示している。すなわち、 1 の一番上のバーと、2 の最初の行と、3 の最初のアライメント結果が対応している。 15 16
© Copyright 2026